So now you don't want to talk about the butterflies?
I understand what I read. You guys don't. I don't know why you think anyone has suggested that the butterfly evolved (PiMPle will cut here) in such a way as YOU described or why you think metamorphosis is evolution, but my guess is both are based on something you read and misunderstood.
Epigenetics is not that hard to understand. It just controls the way genes are read. For instance, methylation can inhibit gene transcription making parts of the DNA unreadable. Butterflies and other species that go through metamorphosis use epigenetics to control the way their genes are read which allows them to unfold the genes into different structures.
What's interesting to me is that the inheritability of the epigenetics might explain a lot about the monarchs and their migration, which is another discussion we've had before.
http://isciencemag.co.uk/blog/butterflies-brains-dna-games/
Since Watson and Crick’s seminal discovery of the double helix there’s been an emergent supposition that DNA makes us what we are. This is a savagely simple belief, which has allowed creationists to pick holes in Darwin’s arguments, but also probably reflects why only 40 years later genetic research is at the point where we can interact with our genomes to provide medical aid.
Most of us have been told we share 70% of our DNA with a banana, 98% with a gorilla and 99% with a chimpanzee. And yet, despite the obvious partiality primates have to eating them, both humans and these species of ape have little in common with bananas. Inevitably you share a gene with the mould you see growing on old bread, or the bacteria causing your skin to break out. When it comes to our genes, it is less about what the genes are, rather when and how they are turned off and on again, and in which combinations. Just in the same way that a research paper and Harry Potter share many words in common, but produce very different text.
Humans have approximately 22,000 genes whilst the simple soy bean has 46,000 and rice 58,000! However our DNA is approximately 10x longer than that of rice; much of it is actually so-called ‘junk DNA’ which contains no genes. Thus despite its deceptively simple appearance the human genome is larger than plants’, if more tightly regulated. This appears to be true for most complex organisms. Thus since discovering the basic structure of genes (in more than one sense), the question has turned to what processes regulate them. These mechanisms are the subject of epigenetic study.
There are two landmark ideas that introduced the idea of epigenetics – literally meaning “around the genes”; an additional method of transferring genetic information.
First is the metamorphosis of a caterpillar to a butterfly. This transformation is incredibly mystical, wonderful and intensely interesting, both as an allegory and a problem-solving exercise. Scientists asked “If DNA explains the production of new species, what happens to genetic information before and after getting into that cocoon?” The answer was nothing. The genes of the caterpillar were identical to those of the butterfly. Creationists had a field day, proclaiming DNA useless, not contributing to an organism’s form. The geneticists were baffled – they knew genes contributed to an organism’s appearance.
The second new idea was that cells from early embryos could become any tissue type; a lung cell or a heart cell for example. But a heart cell would only ever produce heart cells. This specialisation or “differentiation” occurred in development and some people believed it was irreversible, assuming an egg’s DNA intrinsically altered as the foetus developed.
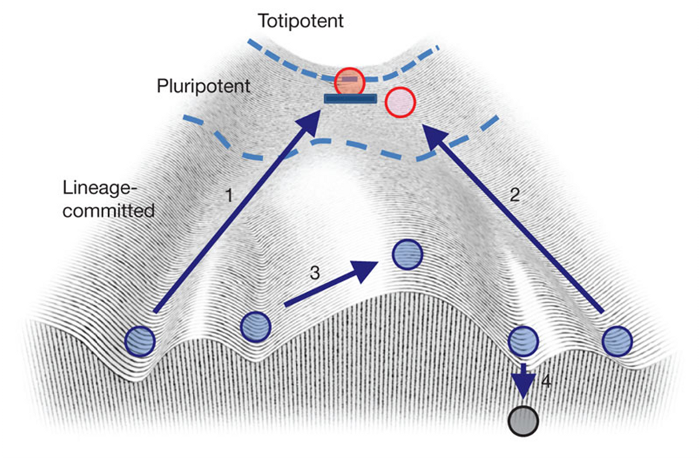
A famous model can be used to explain this – Waddington’s Landscape.
(1) The pathway that reverses a specialised cell to its embryonic stem cell state (2) the pathway reversing a specialised cell to a near embryonic state (3) the route needed to convert one specialist cell directly to another – unlikely (4) cell death
Ignore the circles and arrows for a moment, and consider a slope running downhill towards you with a number of grooves etched into it (there are four on the above). Now place a ball at the top of the slope and let it roll down – it will slot into one of the grooves like a bowling ball into the side gutter. In this analogy, the ball at the top represents something called a totipotent stem cell, capable of becoming any cell type. The second red ball represents a pluripotent stem cell – on that has already started rolling towards the right. The grooves represent cell fates – heart, skin, lung cell etc. The important message is that once a cell is differentiated, it is very difficult to return it to a totipotent state – like rolling the ball back uphill. Rolling a ball from one groove over the large bump in the middle of the landscape is virtually impossible – you have to push the ball back up the trough to the start and roll it back down again.
Thus the theory was that adult cells had already rolled down Waddington’s troughs and these changes were fixed, presumably by some genetic alterations. However Einstein famously said “no amount of experimentation can ever prove me right, but a single experiment can prove me wrong.” Thus the best way of testing this theory was to disprove it.
John Gurdon set about doing exactly this – he took the nuclei of adult frog cells, which contained genetic information, and placed them into frog embryos, replacing the original nucleus. If the adult nucleus was capable of producing an adult frog in this embryonic cell then the idea that adult DNA had no totipotent capacity was wrong. It took many botched attempts but Gurdon eventually grew his impossible frog, and did so again. Hence he demonstrated that there was something about the genetic environment of an adult cell, and not the genes themselves, that prevented totipotency. This heralded the birth of epigenetics; the players around the genes.
What are epigenetic mechanisms?
One of the most common ways of controlling gene expression is the addition of a methyl group to one of the base pairs, cytosine. This tends to occur where cytosine follows guanine, leading to the nickname CpG islands. Where these islands are densely methylated, genes tend to be effectively switched off or ‘silenced’.
Other mechanisms include more global changes. For example DNA is wound round protein balls called histones to organise and condense DNA – each cell contains more than 6 feet of it. These proteins can be chemically altered e.g. by adding a methyl or acetyl group, and once again this affects the activity of local genes.
Furthermore whole chromosomes can be organised across the nucleus to best advantage; gene rich areas tend to cluster centrally where all the transcription machinery is readily available, gene poor areas or repressed genes tend to be excommunicated to the periphery. Loops of DNA are released from their packaging at the border between chromosomes, allowing these to be easily expressed.
It’s become clear that epigenetic regulation is a baffling multiplex process that we’re only just starting to grasp.
Why is it relevant?
Epigenetics also fills in the gaps left by our knowledge of the genome itself – for example why genetically identical twins are two distinctly separate people.
Changes in methylation patterns begin to appear in monozygotic twins even before birth. We know that these changes occur in the last three months. This is intriguing, since it’s also known that a mother’s stress or nutrition status in this critical period can rewire everything from the foetus’s metabolism to its brain.
During WWII the Dutch population starved and researchers used this tragedy to investigate the effect of maternal nutrition on the developing child. Foetal environment had lifelong effects on the infant. Malnutrition in the first three months led to higher incidences of schizophrenia, malnutrition in the latter three months led to the child having reduced lifetime risk of obesity. It’s now known that maternal nutrition alters methylation of foetal DNA, just as dietary content has been shown to induce epigenetic pathways that affect aging. Resveratrol is a compound that locks into this pathway and restores the pattern of methylation seen in young adults; clinical trials have shown efficacy in reducing diabetes and obesity.
It’s clear that these complex, multi-faceted interactions add another dimension to the physical manifestation of genes, representing the interface via which genes and the environment communicate.
Epigenetics explains the metamorphosis of caterpillar to butterfly, foetus to adult, and also shows us ways to reprogram the genetic code, pushing cells back up Waddington’s troughs. The Human Genome Project is complete but now collaborations are underway for the Human Epigenome Project – you can bet that this time you won’t be 70% similar to a banana.
Alcohol reduces methylation of a gene producing the signalling molecule Neuropeptide Y. Neuropeptide release is associated with a reduction in anxiety … epigenetics has perhaps found the rationale for Dutch Courage!
We have already mentioned that methylation changes during development, but it also occurs in aging. One study showed that methylation of genes in the brain increases as we age; effectively more and more genes become silenced. However this can be viewed in two ways. Firstly this gradual dampening of gene expression may be responsible for the general decay of an aging brain, the reduced connections and death of brain cells themselves … or it is a protective mechanism. DNA is vulnerable to mutation – despite extensive proof-reading repair mechanisms. Our repair mechanisms make a mistake at a rate of 1 in 1,000,000,000 … the equivalent of a scribe copying out the bible 280 times before making a single error! However over time these errors will accumulate and may well produce cancer. Damaged genes, when they are irreparable, tend to be switched off. Since we inherit a gene from each parent, we fortunately possess a back-up copy. Thus age-related methylation may well be a way of reducing lethal cancers.
The opposite of gene silencing occurs in schizophrenia – there’s a frenzy of gene expression, methylation utterly haphazard and thinner on the ground, leading to an excited flurry of proteins that ultimately create chaos.
Methylation seems crucial for neural function in particular. This is both tragic and wonderfully hopeful. One psychological disease has the following story; a normal baby girl, meeting all her developmental milestones, happy and chatty until she reaches 2-3 years old. At this point she develops severe mental retardation, loses the ability to speak, draw or interact and won’t even reach for food if placed in front of her. For a time, many people believed this was autism induced by the MMR vaccine, since this unhappily (and non-relatedly) coincides with the age of symptom onset.
However the problem appears to be due to a faulty copy of the gene MeCP2 which helps to read methylated DNA. This gene relies on having the same methylation pattern as the mother to function correctly, whilst some genes require the father’s methylation pattern to behave. However in 2007 a study published an incredible paper showing that this disease, Rett Syndrome, is reversible in mice. The addition of a working copy of MeCP2 reduced or reversed the symptoms. This lends support to the hope that some developmental neurological conditions, which we tend to consider irreversible, may in fact be curable in the future.
Methylation patterns also run amok in cancer and may play a greater role than genes per se in the progression and treatability of this disease.
It’s clear that these complex, multi-faceted interactions add another dimension to the physical manifestation of genes, representing the interface via which genes and the environment communicate.
Epigenetics explains the metamorphosis of caterpillar to butterfly, foetus to adult, and also shows us ways to reprogram the genetic code, pushing cells back up Waddington’s troughs. The Human Genome Project is complete but now collaborations are underway for the Human Epigenome Project – you can bet that this time you won’t be 70% similar to a banana.